Смешанные биополимеры
Давно уже известны четыре класса полимеров живых систем – биополимеров. Это белки, нуклеиновые кислоты, полисахариды и липиды. Четыре, так сказать, «кита».
Как обстоит дело с такими «китами» в естественных науках? Чаще всего так. Сначала – хаотическое нагромождение объектов исследования. Приходит Наблюдатель. Он просто описывает как можно точнее и подробнее отдельные объекты. Мы узнаем много нового, но хаос накопленных фактов остается первозданным. Затем является Систематик. Он группирует факты, объединяет однородные или сходные в классы, типы и т.д., создает некоторую систему поиска – словом, раскладывает все по своим полочкам. На этом этапе обычно и появляются те самые «киты». Затем приходят другие Наблюдатели (чаще всего во множественном числе). Эти говорят: «Позвольте! А вот это не укладывается в вашу систематику, а здесь явно промежуточный тип, а это вообще вне известной серии фактов!». Тогда Систематику (или его последователям) приходится вводить подклассы, подтипы, новые классы и т.п. Наконец, приходит Исследователь и устанавливает, что классы, типы и т.д. – все это хорошо для школьных учебников, а реальные объекты образуют почти непрерывные переходы от одного типа к другому, а сами типы (или классы) суть только схематические описания некоторых крайних, часто нетипичных случаев.
Нуклеиновые кислоты стоят несколько особняком; но для белков, полисахаридов и липидов ситуация сейчас явно близка к той, к которой приходит наш Исследователь. Во многих полисахаридах при ближайшем рассмотрении можно обнаружить большой или маленький ковалентно привязанный пептидный фрагмент. А очень многие классические белки, как выясняется при подробном анализе, несут на поверхности своих глобул короткие олигосахаридные цепи. Однако первые продолжают по инерции называть просто полисахаридами, а вторые – просто белками.
Между этими крайностями имеются всевозможные системы, содержащие больше или меньше белковой компоненты и больше или меньше полисахаридной. Такие соединения называют гликопротеинами, а также протеогликанами (гликаны – общее название полисахаридов). Точного определения у этих терминов нет, и те или иные классы биополимеров называют либо гликопротеинами, либо протеогликанами, руководствуясь при этом скорее традицией, чем какими-либо четкими критериями. Аналогично обстоит дело с ковалентно связанными углеводами и липидами: их называют гликолипидами, а также липополисахаридами. Весь же тип природных высокомолекулярных соединений, включающих ковалентно связанные фрагменты полимеров более чем одного класса, называют смешанными биополимерами, а в последнее время – гликоконъюгатами.
Стурктуры смешанных биополимеров чрезвычайно сложны, а их подробное изучение в сущности лишь только начинается. В отличие от полисахаридов систематически описать и классифицировать типы структур смешанных биополимеров весьма затруднительно прежде всего из-за ограниченного количества надежно и полно расшифрованных структур. Укажем лишь, что связь олиго- или полисахаридной компоненты с пептидной, белковой или липидной осуществляется обычно при помощи гликозидной связи: либо по гидроксильным группам (например, в остатках оксиаминокислот пептидной цепи), либо по амидной группе амидов двухосновных аминокислот. Возможна также фосфодиэфирная связь, подобная той, которая лежит в основе строения нуклеиновых кислот.
Для иллюстрации схематически опишем структуры двух таких биополимеров: гликопротеина и липополисахарида. Биополимеры, определяющие групповую принадлежность ткани, представляют собой высокомолекулярные (молекулярная масса до 1 млн.) гликопротеины, содержащие около 80-85% углеводной компоненты и около 15-20% пептидной. В основе строения их молекул лежит длинная полипептидная цепь с весьма высоким (по сравнению с большинством белков) содержанием оксиаминокислот – серина и треонина.
К гидроксильным группам части этих аминокислотных остатков присоединены гликозидными связями углеводные цепи, общее число которых достигает нескольких сотен. Эти цепи содержат 15-20 моносахаридных остатков каждая, имеют высокоразветвленную структуру и построены из остатков N-ацетил-D-глюкозамина, N-ацетил-D-галактозамина, D-галактозы, L-фукозы, и в части случаев N-ацетил-D-нейраминовой кислоты. В биологическом отношении максимально функциональны (непосредственно ответственны за групповую специфичность) концевые (внешние) участки моносахаридных остатков (так называемые детерминанты). Именно здесь расположены все остатки L-фукозы и N-ацетил-D-нейраминовой кислоты (см. схему).
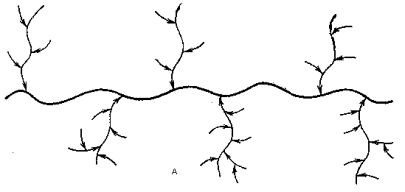
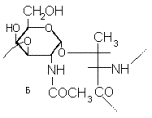
Жирные линии на схеме символизируют полипептидную цепь, остальные линии – полисахаридные цепи: А-общая схема, Б-один из узлов связи полисахаридной и пептидной цепей.
Второй пример – липополисахариды грамотрицательных бактерий, располагающиеся на внешней поверхности бактериальной клетки. На контакт именно с этими биополимерами животный организм-хозяин дает иммунный ответ – начинает вырабатывать антитела. Иными словами, липополисахариды такого типа – это высокоактивные и высокоспецифичные антигены, структура которых строго индивидуальна для каждого вида микроорганизмов. Однако схема построения этих структур имеет весьма общий характер для больших классов микроорганизмов. Вот так приблизительно они построены.
Макромолекула в целом линейна и состоит из трех последовательно связанных крупных блоков, представленных на схеме:
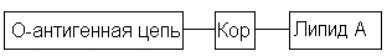
Липид А – главным образом дисахарид, состоящий из двух остатков D-глюкозамина, к одному из гидроксилов которого присоединен кор, а остальные гидроксилы и обе аминогруппы ацилированы высшими жирными кислотами, что и придает фрагменту высоко гидрофобный, липидный характер. Благодаря этому липидный фрагмент липополисахарида погружен (можно было бы сказать, растворен или, еще точнее, вплавлен) в липидную мембрану клетки, что и обеспечивает прочную связь всей молекулы, полисахаридная и, следовательно, высоко гидрофильная, обращена в водную среду, окружающую бактериальную клетку.
Кор представляет собой линейный или слаборазветвленный (по типу гребнеобразного) полисахарид, содержащий остатки довольно необычных моносахаридов – 2-кето-3-дезоксиоктоновых кислот (общая формула 40). Наконец, О-антигенная цепь – это обычно регулярный полисахарид, построенный из повторяющихся три-гексасахаридных (часто разветвленных) звеньев; причем в их состав нередко входят весьма экзотические моносахариды.

Липидная часть и кор сравнительно мало меняются при переходе от одних микроорганизмов к другим в пределах одного класса, тогда как O-антигенная цепь широко варьирует и строго индивидуальна для каждого вида. Как ясно из сказанного, именно эта часть молекулы составляет самый внешний слой бактериальной клетки, с которым непосредственно входит в контакт организм-хозяин при инфекции.
Предыдущая страница |
Следующая страница
СОДЕРЖАНИЕ