Полисахариды
ПОЛИСАХАРИДЫ (гликаны), полимерные углеводы, молекулы к-рых построены из моносахаридных остатков, соединенных гликозидными связями.
Степень полимеризации полисахаридов составляет от 10-20 до неск. тысяч остатков. Каждый моносахаридный остаток в составе полисахаридов может находиться в пиранозной или фуранозной форме и иметь а- или р-конфигурацию гликозидного центра (см. Моносахариды). Моносахаридный остаток способен образовывать одну гликозидную связь с соседним моносахаридом, но может предоставить неск. гидроксильных групп для присоединения др. моносахаридов. В соответствии с этим, как и в случае олигосахаридов, молекулы полисахаридов могут быть линейными или разветвленными. Линейные полисахариды имеют один невосстанавливающий и один восстанавливающий конец; в разветвленных полисахаридах также м. б. только один восстанавливающий конец, тогда как число невосстанавливающих концевых моносахаридных остатков на 1 превышает число разветвлений. Благодаря гликозидной гидроксигруппе восстанавливающего конца молекулы полисахариды могут присоединяться к молекулам неуглеводной природы, напр. к белкам и пептидам с образованием гликопротеинов и протеогликанов, к липидам с образованием липополисахаридов и гликолипидов и т.д.; в сравнительно редких случаях наблюдается образование циклических полисахаридов.
Гидрокси-, карбокси- и аминогруппы моносахаридных остатков, входящих в полисахариды, в свою очередь могут служить местами присоединения неуглеводных группировок, таких, как остатки орг. и неорг. к-т (с образованием ацетатов, сульфатов, фосфатов и др.), пировиноградной к-ты (образующей циклич. ацетали), метанола (образующего сложные эфиры с уроновыми к-тами) и т.д.
Полисахариды, построенные из остатков только одного моносахарида, наз. гомополисахаридами (гомогликанами); в соответствии с природой этого моносахарида различают глю-каны, маннаны, галактаны, ксиланы, арабинаны и др. Полное название полисахарида должно содержать информацию об абс. конфигурации входящих в его состав моносахаридных остатков, размере циклов, положении связей и конфигурации гликозидных центров; в соответствии с этими требованиями строгим назв., напр., целлюлозы будет поли(1 : 4)-b-D-глюкопиранан.
Полисахариды, построенные из остатков двух и более моносахаридов, наз. гетерополисахаридами (гетерогликанами). К ним относятся глюкоманнаны, арабиногалактаны, араби-ноксиланы и др. Строгие назв. гетерогликанов (а также и гомополисахаридов, содержащих разветвления или неск. типов связей) громоздки и неудобны в употреблении; обычно пользуются широко распространенными тривиальными назв. (напр., гепарин, гликоген, инулин, ламтаран, хитин), а для изображения структурных ф-л часто применяют сокращенную запись (см. также Олигосахариды):
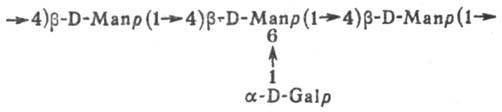
Галактоманнан; a-D-галактопирано-b-D-маннопиранан (Manp и Galp- соотв. остатки маннозы и галактозы в пиранозной форме)
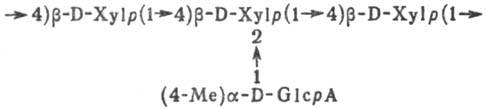
4-О-Метилглюкуроноксилан; (4-О-метил)-a-D-глюкопиран-уроно-b-D-ксилопиранан (Xylp и GlcpA-соотв. остатки ксилозы и глюкуроновой к-ты в пиранозной форме, Me = СН3)
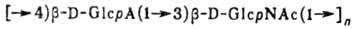
Гиалуроновая к-та, глюкозаминоглюкуроногликан; 2-ацет-амидо-2-дезокси-b-D-глюкопирано-b-D-глюкопирануроно-гликан [Ас = СН3С(О)]
Полисахариды в природе составляют главную массу орг. в-ва, находящегося в биосфере Земли. Они выполняют в живых организмах три важнейших типа биол. ф-ций, выступая в роли энергетич. резерва, структурных компонентов клеток и тканей или же защитных в-в.
Хорошо известными резервными полисахаридами являются крахмал, гликоген, фруктаны, галактоманнаны и нек-рые р-глюканы. Эти полисахариды способны быстро гидролизоваться имеющимися в клетках ферментами, и их содержание сильно зависит от условий существования и стадии развития организма.
Структурные полисахариды можно разделить на два класса. К первому относят нерастворимые в воде полимеры, образующие волокнистые структуры и служащие армирующим материалом клеточной стенки (целлюлоза высших растений и нек-рых водорослей, хитин грибов, b-D-ксиланы и b-D-ман-наны нек-рых водорослей и высших растений). Ко второму классу относят гелеобразующие полисахариды, обеспечивающие эластичность клеточных стенок и адгезию клеток в тканях. Характерными представителями этого класса полисахаридов являются сульфатир. гликозаминогликаны (мукополисахариды) соединит. ткани животных, сульфатир. галактаны красных водорослей, альгиновые к-ты, пектины и нек-рые гемицеллюло-зы высших растений.
К защитным полисахаридам относят камеди высших растений (гетеро-полисахариды сложного состава и строения), образующиеся в ответ на повреждение растит. тканей, и многочисл. внеклеточные полисахариды микроорганизмов и водорослей, образующие защитную капсулу или модифицирующие св-ва среды обитания клеток.
Биосинтез полисахаридов. Все разнообразие структур природных полисахаридов-результат трех типов биосинтетич. процессов. Первым из них служит последоват. перенос отдельных моносахаридных остатков от нуклеотидсахаров на растущую цепь с участием специфич. ферментов гликозилтрансфераз, обеспечивающих необходимое положение и стереохимию образующейся гли-козидной связи; таким способом синтезируются как моно тонные последовательности моносахаридных остатков в го могликанах, так и лишенные признаков регулярности гете-рополисахаридные цепи гликопротеинов.
Второй тип-сборка олигосахаридного "повторяющегося звена" по первому типу р-ций и его последующая полимеризация с образованием строго регулярных полимерных молекул, характерных для полисахаридных цепей липополисахаридов грамотрицательных бактерий или для бактериальных капсульных полисахаридов.
Наконец, полисахариды, построенные по первому или второму типу, могут испытывать постполимеризац. модификации (третий тип биосинтеза), к-рые включают замещение атомов Н гид-роксильных групп на ацильные остатки (ацетилирование, сульфатирование), присоединение боковых моно- и олигоса-харидных остатков и даже изменение конфигурации отдельных моносахаридных звеньев [таким путем в результате эпимеризации при атоме С-5 образуются остатки L-гулуро-новой к-ты из D-маннуроновой в составе альгинатов (см. Альгиновые кислоты), а также остатки L-идуроновой к-ты из D-глюкуроновой в составе мукополисахаридов]. Последние р-ции часто приводят к нарушению (маскировке) первонач. регулярности цепей полисахаридов и к образованию нерегулярных (мн. гемицеллюлозы) или блочных (альгиновые к-ты, мукополисахариды) структур.
Свойства. Большинство полисахаридов-бесцв. аморфные порошки, разлагающиеся при нагр. выше 200 °С. Полисахариды, молекулы к-рых обладают разветвленной структурой или имеют полианионный характер благодаря карбоксильным или сульфатным группам, как правило, достаточно легко раств. в воде, несмотря на высокие мол. массы, тогда как линейные полисахариды, обладающие жесткими вытянутыми молекулами (целлюлоза, хитин), образуют прочные упорядоченные надмолекулярные ассоциаты, в результате чего практически не раств. в воде. Известны промежут. случаи блочных молекул полисахаридов, в к-рых одни участки склонны к межмол. ассоциации, а другие-нет; водные р-ры таких полисахаридов при определенных условиях переходят в гели (пектины, альгиновые к-ты, кар-рагинаны, агар).
Р-римые полисахариды можно осадить из водных р-ров смешивающимися с водой орг. р-рителями (напр., этанолом, метанолом, ацетоном). Р-римость конкретного полисахарида определяет методику выделения его из прир. объекта. Так, целлюлозу и хитин получают, отмывая подходящими реагентами все сопутствующие в-ва, тогда как прочие полисахариды вначале переводят в р-р и выделяют затем фракционным осаждением р-рителями, с помощью образования нерастворимых комплексов или солей, ионообменной хроматографией и т.д.
Солюбилизация сложных надмолекулярных комплексов (напр., полисахариды клеточных стенок) требует подчас достаточно жестких условий, не исключающих расщепления нек-рых хим. связей. Выделенные полисахаридные препараты обычно представляют собой смеси полимергомологичных молекул; в случае нерегулярных полисахаридов дополнит. фактором неоднородности служит т. наз. микрогетерогенность-различия отдельных молекул друг от друга по степени протекания постполимеризац. модификаций.
Из хим. р-ций полисахаридов важное значение имеет гидролиз гликозидных связей под действием разб. минер. к-т, позволяющий получить моносахариды, входящие в состав полисахаридов. В отличие от олигосахаридов, восстанавливающие св-ва или мутаротация (связанные с наличием в молекуле концевой карбонильной группы) в полисахаридах проявляются слабо из-за их больших мол. масс. Наличие множества гидроксильных групп позволяет проводить р-ции алкилирования или ацилирования; нек-рые из них имеют существ. значение для установления строения или практич. использования полисахаридов.
Установление строения. Установление первичной структуры полисахарида складывается из последоват. решения трех задач: определения состава, типов связей между моносахаридами и последовательности отдельных моносахаридных звеньев. Первая задача решается гидролизом и количеств. определением (одним из видов количеств. хроматографии, а в отдельных случаях-с помощью фотоколориметрии) всех входящих в состав полисахарида моносахаридов, а также неуглеводных заместителей (если они имеются).
Для определения типов связей между моносахаридами обычно служит метод метилирования, к-рый заключается в превращении всех своб. гидроксильных групп полисахаридов в метиловые эфиры. Поскольку эти группировки устойчивы в уcловиях кислотного гидролиза гликозидных связей, то гидролиз метилированного полисахарида дает набор метиловых эфиров моносахаридов. Они различаются числом групп СН3 в зависимости от положения-моносахаридного остатка в полимерной молекуле. Так, концевые невосстанавливающие остатки гексоз дают тетра-О-метилпроизводные, остатки гексоз из линейных участков цепей-три-О-метилпроизводные, из точек разветвления-ди-О-метилпроизводные и т.д. Наличие своб. гидроксильных групп в метилированных моносахаридах обусловлено тем, что в родоначальном полисахариде эти гидроксилы участвовали в образовании либо циклич. фо'рм моносахаридов (пиранозных или фуранозных), либо гликозидных связей. Поэтому определение положения групп СН3 (а следовательно, и гидроксильных) в каждом таком производном позволяет в принципе установить размер цикла родоначаль-ного моносахаридного остатка в полимерной цепи и место замещения его соседним моносахаридным остатком (или остатками).
Существующие методики метилирования полисахаридов (напр., метод Хакомори- действие NaH в ДМСО и затем СН3I) обладают весьма высокой эффективностью и пригодны для микроколичеств в-ва. Анализ продуктов метилирования проводится с применением хромато-масс-спектрометрии и дает надежные сведения о положении групп СН3 в производных моносахаридов.
Сведения о конфигурации гликозидных центров и последовательности моносахаридных остатков в полимере получают, проводя частичное расщепление молекул полисахаридов и устанавливая строение образующихся при этом олигосахаридов. Универсальным методом расщепления является частичный кислотный гидролиз, однако в общем случае он дает сложные смеси олигосахаридов с небольшими выходами. Лучшие результаты получаются при более специфич. воздействии на молекулу полисахарида хим. реагентами (ацетолиз, сольволиз безводным HF) или ферментами.
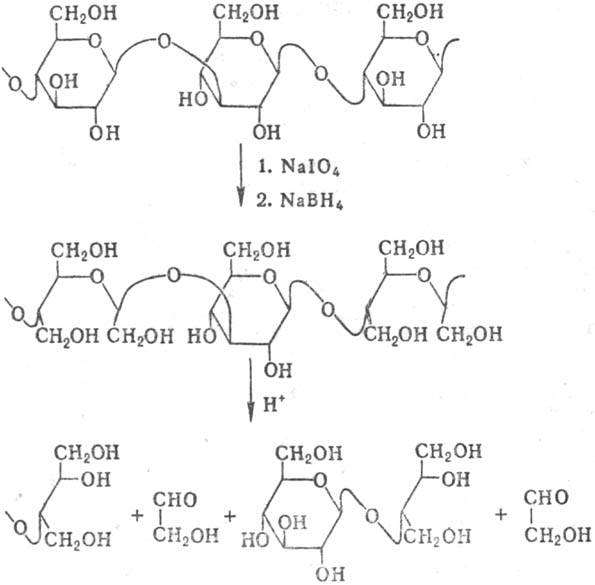
Своеобразный способ фрагментации молекул полисахаридов-расщепление по Смиту, включающее периодатное окисление, восстановление полученного полиальдегида в полиол действием NaBH4 и мягкий кислотный гидролиз, разрушающий ацетальные группировки (но не гликозидные связи моносахаридов, не затронутых периодатным окислением). Метод Смита часто позволяет получить фрагменты молекул полисахаридов, недоступные при обычном кислотном или ферментативном гидролизе (стадия образования полиальдегидов не показана):
С хим. методами установления первичной структуры полисахаридов успешно конкурирует спектроскопия ЯМР. Спектры ПМР и ЯМР13С содержат ценнейшую информацию о функцион. составе полисахаридов, положениях межмономерных связей, размерах циклов моносахаридных остатков, конфигурациях гликозидных центров и последовательности моносахаридов в цепи; из спектров ЯМР13С можно определить абс. конфигурации отдельных моносахаридных остатков (если известны абс. конфигурации соседних звеньев), а также получить данные о регулярном строении полисахаридов. Если известен моносахаридный состав линейного регулярного полисахарида, построенного из повторяющихся олигосахаридных звеньев, то задача установления его полного строения по спектру ЯМР успешно решается с помощью соответствующих компьютерных программ.
Др. физ.-хим. методы исследования применяются для определения мол. масс полисахаридов (вискозиметрия, светорассеяние, ультрацентрифугирование) и конформации молекул в твердом состоянии (рентгенография напряженных волокон или пленок).
Синтез полисахаридов. Синтез природных полисахаридов и их аналогов представляет интерес для установления связи их строения и биол. активности, в первую очередь иммунологич. св-в бактериальных полисахаридов.
Поликонденсация моносахаридов под действием кислых катализаторов приводит к полимерным продуктам, содержащим хаотич. набор межмономерных связей, катионная полимеризация защищенных 1,6-ангидридов гексоз-к линейным 1,6-связанным полисахаридам. Для общего решения задачи направленного синтеза сложных природных полисахаридов необходимы методы стереоспецифич. гликозилирования, пригодные для полимеризации или поликонденсации олигосахаридов.
Примером Такой р-ции служит взаимод. цианоэтилидено-вых производных углеводов с тритиловыми эфирами Сахаров, приводящее к 1,2-транс-гликозидам:
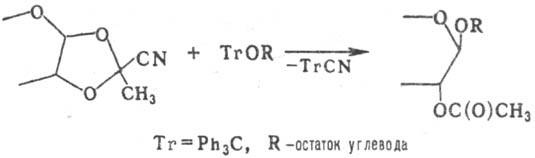
Поскольку обе группировки (тритиловую и цианоэтилидено-вую) можно ввести в одну молекулу моно- или олигосахари-да, поликонденсация такого производного приводит к полисахаридам заданного строения. Этим путем были синтезированы полисахариды, содержащие ди-, три- и тетрасахаридные повторяющиеся звенья, в т.ч. идентичные природным полисахаридам бактериального происхождения.
Др. перспективный подход к синтезу полисахаридов-химико-ферментативный метод, в к-ром наиб. трудные стадии получения олигосахаридных предшественников или их полимеризация проводятся с использованием соответствующих ферментов. Показано, что этим путем можно получать не только природные полисахариды, но и их аналоги; недостатком метода является сравнительно малая доступность необходимых ферментов.
Применение. Многие полисахариды производят в крупных масштабах, они находят разнообразное практич. применение. Так, целлюлозу используют для произ-ва бумаги и искусств. волокна, целлюлозы ацетаты - для волокон и пленок, целлюлозы нитраты - для ВВ, а водорастворимые метилцеллюлозу гидроксиэтилцеллюлозу и карбоксиметилцеллюлозу - как стабилизаторы суспензий и эмульсий.
Крахмал используют в пищ. пром-сти, где находят применение в качестве текстурир. агентов также пектины, альгина-ты, каррагинаны и галактоманнаны. Перечисленные полисахариды имеют растит. происхождение, но с ними все успешнее конкурируют бактериальные полисахариды, получаемые в результате пром. микробиол. синтеза (ксантан, образующий стабильные высоковязкие р-ры, и другие полисахариды со сходными св-вами).
Весьма перспективны разнообразные техн. применения хитозана (кагионного полисахарида, получаемого в результате дезаце-тилирования прир. хитина).
Многие полисахариды применяют в медицине (агар в микробиологии, гидроксиэтилированный крахмал и декстраны в качестве плазмозамещающих р-ров, гепарин как антикоагулянт, нек-рые глюканы грибов как противоопухолевые и иммуностимулирующие агенты), биотехнологии (альгинаты и каррагинаны как среда для иммобилизации клеток) и лаб. технике (целлюлоза, агароза и их производные как носители при разл. способах хроматографии и электрофореза).
Лит.: Химия углеводов, М., 1967, с. 477-624; Прогресс химии углеводов, М., 1985; The polusaccharides, ed. by G.O. Aspinall, v. 1-3, N.Y., 1982-85.
А.И. Усов.