Цикл мочевины
Деградация аминокислот происходит преимущественно в печени. При этом непосредственно или косвенно освобождается аммиак. Значительные количества аммиака образуются при распаде пуринов и пиримидинов.
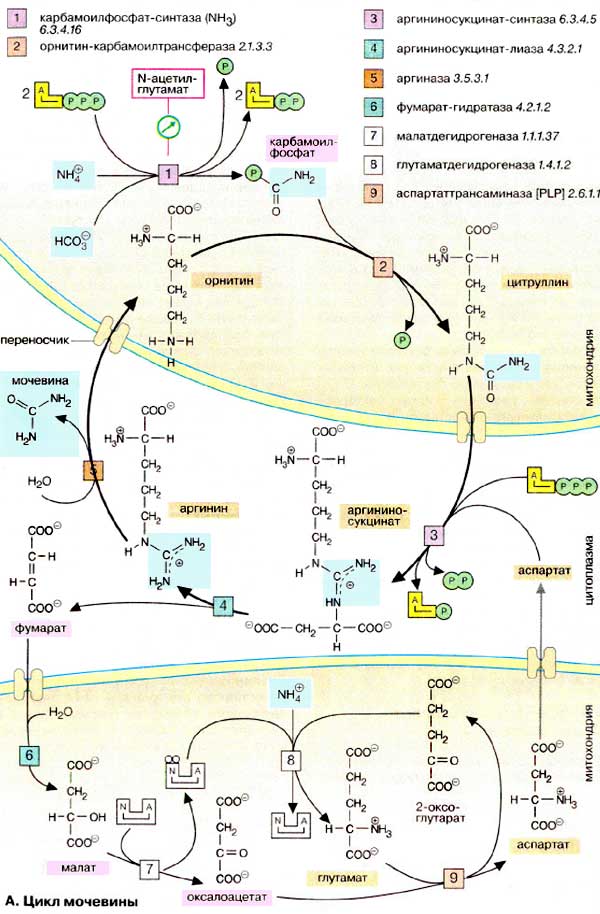
Аммиак (на схеме наверху слева), основание средней силы, является клеточным ядом. При высоких концентрациях он повреждает главным образом нервные клетки. Поэтому аммиак должен быстро инактивироваться и выводиться из организма. В организме человека это осуществляется прежде всего за счет образования мочевины (на схеме в середине слева), часть NH3 выводится непосредственно почками (см. рис. 319).
У разных видов позвоночных инактивация и выведение аммиака производятся различными способами. Живущие в воде животные выделяют аммиак непосредственно а воду; например, у рыб он выводится через жабры (аммониотелические организмы). Наземные позвоночные, в том числе человек, выделяют лишь небольшое количество аммиака, а основная его часть превращается в мочевину (уреотелические организмы). Птицы и рептилии, напротив, образуют мочевую кислоту, которая в связи с экономией воды выделяется преимущественно в твердом виде (урикотелические организмы).
А. Цикл мочевины
Мочевина является диамидом угольной кислоты. В противоположность аммиаку это нейтральное и нетоксичное соединение. При необходимости небольшая молекула мочевины может проходить через мембраны. По этой причине, а также из-за ее хорошей растворимости в воде мочевина легко переносится кровью и выводится с мочой.
Мочевина образуется в результате циклической последовательности реакций, протекающих в печени. Оба атома азота берутся из свободного аммиака и за счет дезаминирования аспартата, карбонильная группа — из гидрокарбоната. На первой стадии, реакция [1], из гидрокарбоната (НСО3-) и аммиака с потреблением 2 молекул АТФ образуется карбамоилфосфат. Как ангидрид это соединение обладает высоким реакционным потенциалом. На следующей стадии, реакция [2], карбамоильный остаток переносится на орнитин с образованием цитруллина. Вторая аминогруппа молекулы мочевины поставляется за счет реакции аспартата (на схеме внизу справа) с цитруллином [3]. Для этой реакции вновь необходима энергия в форме АТФ, который при этом расщепляется на АМФ и дифосфат. Для обеспечения необратимости реакции дифосфат гидролизуется полностью (не показано). Отщепление фумарата от аргининосукцината приводит к аргинину [4], из которого в результате гидролиза образуется изомочевина [5], сразу же превращающаяся в результате перегруппировки в мочевину. Остающийся орнитин вновь включается в цикл мочевины.
Фумарат, образующийся в цикле мочевины, может в результате двух стадий цитратного цикла [6, 7] через малат переходить в оксалоацетат, который за счет трансаминирования [9] далее прекращается в аспартат. Последний также вновь вовлекается в цикл мочевины.
Биосинтез мочевины требует больших затрат энергии. Необходимая энергия поставляется за счет расщепления четырех высокоэнергетических связей: двух при синтезе карбамоилфосфата и двух (!) при образовании аргининосукцината (АТФ → АМФ + PPi, РРi → 2Pi).
Цикл мочевины протекает исключительно в печени. Он разделен на два компартмента, митохондрии и цитоплазму. Прохождение через мембрану промежуточных соединений цитруллина и орнитина возможно только с помощью переносчиков. Обе аминокислоты небелкового происхождения.
Скорость синтеза мочевины определяется первой реакцией цикла [1]. Карбамоилфосфатсинтаза активна только в присутствии N-ацетилглутамата. Состояние обмена веществ (уровень аргинина, энергоснабжение) сильно зависит от концентрации этого аллостерического эффектора.