Первичная структура нуклеиновых кислот
Под первичной структурой нуклеиновых кислот понимают порядок, последовательность расположения мононуклеотидов в полинуклеотидной цепи ДНК и РНК. Такая цепь стабилизируется 3',5'-фосфодиэфирными связями. Поскольку молекулярная масса нуклеиновых кислот колеблется в широких пределах (от 2•104 до 1010–1011), установить первичную структуру всех известных РНК и особенно ДНК весьма сложно. Тем не менее во всех нуклеиновых кислотах (точнее, в одноцепочечной нуклеиновой кислоте) имеется один и тот же тип связи – 3',5'-фосфодиэфирная связь между соседними нуклеотидами. Эту общую основу структуры можно представить следующим образом:
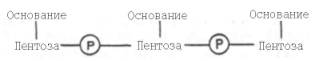
Установлено, что в образовании межнуклеотидной связи участвуют гидроксильные группы в 3'- и 5'-положениях остатков углевода.
К настоящему времени удалось определить первичную структуру почти всех тРНК, ряда молекул 5S рРНК, 16S рРНК E.coli, вирусных РНК, в состав которых входят сотни и тысячи нуклеотидных остатков. Ниже приводится примерная схема последовательности нуклеотидов в молекуле РНК. Все клеточные РНК в основном состоят из одноцепочечной по-линуклеотидной цепи:
5'-Г–У–Г–Ц–А–А–...–У–Ц–Г–Ц–Ц–А–3'
Полинуклеотидная цепь молекулы РНК имеет на одном конце почти всегда свободный монофосфорный эфир, который принято обозначать как 5'-конец; на противоположном конце цепи такой фосфат отсутствует, а содержится нуклеотид со свободными 2'- и 3'-гидроксильными группами. Если подвергнуть щелочному гидролизу молекулу РНК, то в качестве концевого нуклеотида будут обнаружены ЦМФ со свободным фосфатом у 5'-конца и свободный аденозин в виде свободного нуклеозида у 3'-конца полинуклеотидной цепи.
В выяснении первичной структуры РНК решающую роль сыграли методы ступенчатого гидролиза, осуществленного в основном экзонуклеа-зами и заключающегося в последовательном отщеплении по одному мононуклеотиду с одного конца молекулы нуклеиновой кислоты. Ниже представлена первичная структура первой РНК, имеющей 77 нуклеотидов, для которой была расшифрована нуклеотидная последовательность в 1965 г. Р. Холли и сотр., а именно аланиновой тРНК:
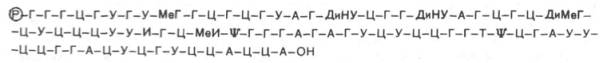
В этой структуре Р – остаток фосфата, ψ – псевдоУМФ, МеГ – метилгуа-нин, ДиНУ – дигидроурацил, ДиМеГ – диметилгуанин, МеИ – метилинозин.
Следует особо указать на две существенные особенности первичной структуры всех тРНК. Первая из них заключается в том, что 5'-концом всегда является гуаниловая (редко цитидиловая) кислота, несущая свободный остаток фосфата у С-5'. Вторая особенность – наличие на противоположном конце молекулы остатков трех мононуклеотидов с одинаковой последовательностью – ЦЦА, причем остаток адениловой кислоты содержит свободную 3'-ОН-группу.
Между этими структурами в строго определенной последовательности располагаются все остальные нуклеотидные остатки, среди которых на долю минорных нуклеотидов приходится до 10%. Полинуклеотидная цепь разных типов тРНК содержит около 75 нуклеотидов.
Матричные (информационные) РНК относятся к наиболее гетерогенному классу нуклеиновых кислот, отличающихся по массе (см. табл. 3.1), структуре, размерам, стабильности и функциям. Основной функцией мРНК является перенос информации от ДНК (точнее, от гена) на белоксинте-зирующую систему клетки. мРНК выполняет роль матрицы и, следовательно, определяет первичную структуру синтезируемого белка (подробнее см. главу 14). мРНК наделены рядом особенностей первичной структуры; в частности, на 5'-конце все они содержат определенную последовательность рибонуклеотидов, получившую название шапочки (кэп). Первым нуклеотидом является 7-метилгуанозинтрифосфат, который присоединяется к 5'-гидроксилу соседнего мононуклеотида, представленного 2'-О-метилпуриновым нуклеотидом. На другом 3'-конце большинства (но не всех) мРНК содержится полиадениловая последовательность (поли-А), насчитывающая от 150 до 200 нуклеотидов.
Роль «кэпирования» и «полиаденилирования» мРНК в белковом синтезе окончательно не выяснена. Предполагают, что кэп необходим для специфического узнавания в процессе трансляции, в то время как поли-А отводится роль фактора стабилизации всей молекулы мРНК.
В последние годы расшифрована первичная структура не только низкомолекулярных 5S рРНК разных бактерий и 5,8S рРНК клеток животных, но и высокомолекулярных 16S и 18S рРНК, насчитывающих до 1200–1500 нуклеотидных звеньев. Более того, уже выяснены нуклеотидные последовательности 23S рРНК E.coli и 25S рРНК дрожжевой клетки, а также первичные структуры высокомолекулярных (28S) рРНК клеток эукариот, насчитывающих около 4700 нуклеотидов.
В настоящее время проводятся исследования первичных структур различных молекул ДНК. Около 15 лет назад была полностью расшифрована нуклеотидная последовательность митохондриальной ДНК человека (16569 пар нуклеотидов). Известны полные нуклеотидные последовательности ДНК ряда вирусов и плазмид. Совсем недавно завершено определение нуклеотидных последовательностей геномов двух прокариотических организмов (Haemophilus influenzae и Mycoplasma genitalum) и появились сообщения о расшифровке генома первого эукариотического организма – дрожжей. Близки к завершению аналогичные исследования генома E.coli и генома нематоды Caenorhabditis elegans. Исследователи активно работают над полной расшифровкой генома человека.
Результаты секвенирования (определение нуклеотидной последовательности) разных молекул ДНК накапливаются в виде компьютерных банков данных, которые уже доступны для пользователей международных компьютерных сетей (например, «Internet»). Ниже представлены три варианта схемы нуклеотидной последовательности ДНК:
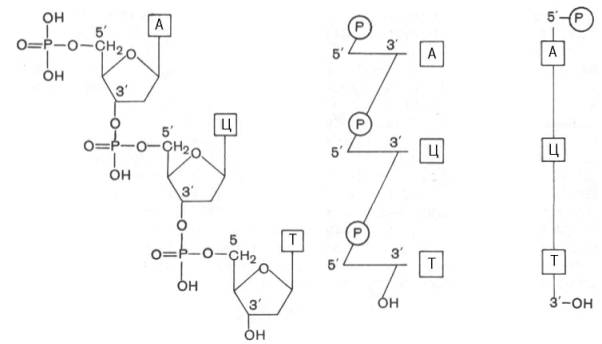
В последнее время о первичной структуре ДНК (точнее, отдельных ее фрагментов) судят по ряду косвенных данных, например, по степени сплоченности нуклеотидных звеньев в молекуле ДНК (определение сводится в конечном счете к выяснению числа и структуры отдельных фракций нуклеотидов, так называемых изоплитов), также по кинетике реассоциации ДНК (метод позволяет выяснить наличие в молекуле повторяющихся последовательностей нуклеотидов). О первичной структуре ДНК судят, кроме того, по распределению минорных оснований (имеются данные о существовании подобной закономерности) и обнаружению в ДНК и определению последовательности палиндромов («обратно бегущие» последовательности, или перевертыши), которые обнаруживаются главным образом в местах рестрикции (см. главу 13). Большие надежды в определении первичной структуры ДНК исследователи возлагают на физические, химические (синтез генов), генетические и другие методы, а также на методы выделения некоторых генов (или их фрагментов) из природных источников и синтеза генов на мРНК при участии фермента обратной транскриптазы. Для установления первичной структуры ДНК недавно предложен экспресс-метод, включающий применение двух ДНК-полимераз (из E.coli и из бактериофага Т4). Однако во всех этих случаях определяется структура небольшого участка ДНК, поэтому полная расшифровка первичной структуры ДНК генома человека ждет своего решения.