Рибосомы: инициация трансляции
Биосинтез белка (трансляция), как и активация аминокислот, происходит в цитоплазме. Он осуществляется нуклеопротеидными частицами, называемыми рибосомами.
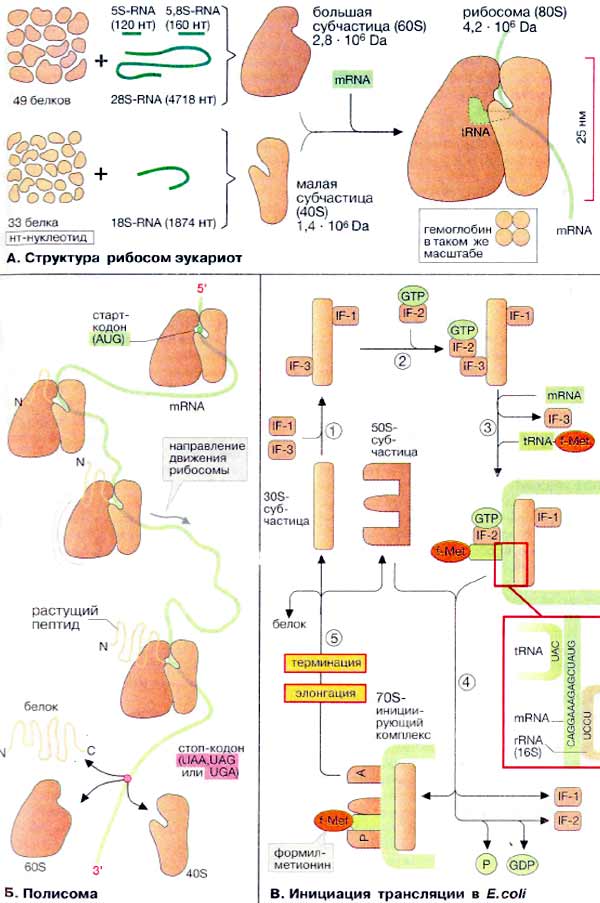
А. Структура рибосом эукариот
Рибосомы состоят из двух различных субчастиц, каждая из которых построена из рибосомной РНК [рРНК (rRNA)] и многих белков. Рибосомы и их субчастицы обычно классифицируют не по массам, а в соответствии с коэффициентами седиментации. Так. коэффициент седиментации полной эукариотической рибосомы составляет около 80 единиц Сведберга (80S), а коэффициент седиментации ее субчастиц составляет 40S и 60S.
Меньшая 40S-субчастица состоит из одной молекулы 18S-рРНК и 30-40 белковых молекул. Большая 60S-субчастица содержит три типа рРНК с коэффициентами седиментации 5S, 5,8S и 28S и 40-50 белков (например, рибосомы гепатоцитов крысы включают 49 белков). В присутствии мРНК (mRNA) субчастицы объединяются с образованием полной рибосомы, масса которой примерно в 650 раз больше массы молекулы гемоглобина. Рибосомы имеют диаметр 20-200 нм и их можно видеть в электронный микроскоп. Структурная организация рибосом полностью не выяснена. Однако известно, что молекулы мРНК проходит через щель около характерной структуры в виде «рога» на малой субчастице, причем эта щель ориентирована как раз в промежуток между двумя субчастицами. тРНК также связываются вблизи этого участка. Для сравнения на схеме в том же масштабе показана молекула тРНК.
Рибосомы прокариот имеют аналогичную структуру, но они несколько мельче, чем эукариотические (коэффициенты седиментации полной рибосомы 70S, а субчастиц — 30S и 50S). Рибосомы митохондрий и хлоропластов близки к прокариотическим.
Б. Полисома
Клетки, в которых происходит активный синтез белков, часто содержат рибосомы, расположенные одна за другой подобно жемчужинам на нитке, в виде так называемой полисомы. Это объясняется тем, что одна молекула мРНК может транслироваться одновременно несколькими рибосомами. Первой стадией трансляции является связывание рибосомы со стартовым (инициирующим) кодоном (AUG, см. с. 244) вблизи 5'-конца мРНК. По мере трансляции рибосома движется по направлению к 3'-концу до тех пор, пока не дойдет до терминирующего кодона (стоп-кодона) (UAA, UAG или UGA). Как только рибосома достигает стоп-кодона, происходит освобождение синтезированного белка и диссоциация рибосомы на отдельные субчастицы.
В. Инициация трансляции в E.coli
Синтез белка у прокариот в основном аналогичен синтезу у эукариот. Здесь и на с. 248 процесс трансляции обсуждается на примере бактерии Escherichia coli.
Первую фазу трансляции — инициацию — можно разделить на несколько стадий. На первой стадии два белковых фактора инициации IF-1 и IF-3 связываются с 30S-субчастицей (1). Затем еще один белковый фактор, IF-2, образует комплекс с ГТФ (GTP) (2), что облегчает ассоциацию 30S-субчастицы с мРНК (mRNA) и связывание тРНК (tRNA), соответствующей инициирующему кодону (3). У прокариот стартовая тРНК несет N-формилметионин (f-Met), у эукариот — метионин. В завершение 50S-субчастица связывается с вышеупомянутым комплексом (4). На третьей и четвертой стадиях идет освобождение факторов инициации и гидролиз связанного с IF-2 ГТФ до ГДФ (GDP) и неорганического фосфата Р. Таким образом, связанный с 70S-рибосомой инициирующий комплекс содержит формилметионин-тРНК в тРНК-связывающем участке, называемом пептидильным участком (Р). Второе место связывания, акцепторный участок (А), во время этой фазы трансляции остается свободным.