Биосинтез ДНК
Прежде чем изложить современные представления о механизме биосинтеза ДНК, следует представить сведения о синтезе этого соединения в бес-
клеточной системе, которыми располагает биохимия. Известно, что для любого синтеза полимерной органической молекулы, осуществляемого in vitro или in vivo, требуется энергия. Источником энергии в реакциях полимеризации мононуклеотидов является энергия, освобождаемая всеми четырьмя типами дезоксирибонуклеозидтрифосфатов, участвующих в синтезе ДНК. Образующийся пирофосфат под действием пирофосфатазы также расщепляется на две молекулы ортофосфата, давая дополнительную энергию для биосинтеза ДНК.
Помимо энергии, биогенез ДНК требует наличия специфических ферментов, катализирующих отдельные этапы синтеза, и множества белковых факторов, абсолютно необходимых для регулирования процесса репликации и проявления каталитической активности ферментов.
Ферментные системы синтеза ДНК у про- и эукариот до конца не выяснены. По имеющимся данным, в репликации ДНК, включающей узнавание точки начала процесса, расплетение родительских цепей ДНК в репликационной вилке, инициацию биосинтеза дочерних цепей и дальнейшую их элонгацию и, наконец, окончание (терминация) процесса, участвует более 40 ферментов и белковых факторов, объединенных в единую ДНК-репликазную систему, называемую реплисомой.
После открытия в 1958 г. А. Корнбергом у Е. coli фермента, катализирующего биосинтез ДНК и названного ДНК-полимеразой I, в течение почти 10 лет считалось, что этот фермент является единственной по-лимеразой, принимающей участие в репликации ДНК in vitro . Однако позже был открыт мутант Е. coli, лишенный ДНК-полимеразы I, но способный синтезировать ДНК с нормальной скоростью. Оказалось, что для репликации ДНК Е. coli необходимо участие нескольких ферментов. ДНК-полимераза I не наделена способностью инициировать синтез цепей ДНК de novo. Одним из хорошо изученных ферментов, участвующих в стадии инициации репликации ДНК, является специфическая клеточная РНК-полимераза, названная праймазой, которая катализирует синтез короткого олигорибонуклеотида (от 10 до 60 нуклеотидов), т.е. праймера, с которого затем начинается синтез ДНК. Праймазы различаются как по структуре, так и по специфичности действия. Получены новые данные о существенной роли праймасомы в каталитическом действии фермента. Праймасома представлена ансамблем из 7 различных субъединиц, включающих около 20 полипептидов общей мол. массой 70000. При помощи белка n' праймасома подвергается быстрому перемещению к отстающей цепи ДНК за счет энергии, генерируемой АТФазной активностью белка n'. В состав праймасомы входит также комплекс белков dna В и dna С, который вблизи репликационной вилки периодически участвует в формировании специфической вторичной структуры ДНК, подходящей для узнавания праймазой.
Основным ферментом, катализирующим биосинтез новообразованной ДНК (точнее, стадию элонгации репликации ДНК), является ДНК-поли-мераза III, представляющая собой мультимерный комплекс собственно ДНК-полимеразы (мол. масса около 900000) и ряда других белков. ДНК-полимераза III из Е. coli состоит минимум из 10 субъединиц. Одна из них – β-субъединица получена в кристаллическом виде, и выяснена ее третичная структура. Имеются доказательства, что в димерной форме ДНК-полимераза III катализирует сопряженный синтез ведущей (лидирующей) и отстающей цепей ДНК при репликации (см. далее). Более точно выяснена также роль ДНК-полимеразы I: она катализирует отщепление затравочного олигорибонуклеотидного праймера и заполнение образующихся после этого пробелов (ниш) дезоксирибонуклеотидами. Известно, что ДНК-полимеразы II из Е. coli (мол. масса 88000) выполняет «ремонтные» функции, исправляя повреждения цепей ДНК. Укажем также, что ДНК-полимераза I в качестве матрицы использует одноцепочечные участки, в то время как ДНК-полимераза III – двухцепочечные ДНК, в которых имеются короткие одноцепочечные последовательности.
Важную функцию соединения двух цепей ДНК или замыкания двух концов одной цепи ДНК в процессе репликации либо репарации ДНК выполняет особый фермент – ДНК - лигаза, катализирующая за счет энергии АТФ образование фосфодиэфирной связи между 3'-ОН-группой де-зоксирибозы одной цепи и 5'-фосфатной группой другой цепи ДНК.
Функцию раскручивания (расплетения) двойной спирали ДНК в репли-кационной вилке, происходящего за счет энергии гидролиза АТФ, выполняет специфический rep-белок, названный хеликазой (мол. масса 300000). Образовавшиеся на определенное время одноцепочечные участки ДНК служат в качестве матрицы при репликации и стабилизируются при помощи особых белков, связывающихся с одноцепочечной ДНК (ДНК-связывающие белки) и препятствующих обратному комплементарному взаимодействию цепей ДНК (мол. масса 75600). В связи с этим их иногда называют дестабилизирующими двойную спираль белками. Имеются, кроме того, особые ферменты топоизомеразы (у прокариот одна из них названа ДНК-гиразой), которые играют особую роль в сверхспирализации, обеспечивая как репликацию, так и транскрипцию ДНК. Эти ферменты наделены способностью не только создавать супервитки, но и уничтожать суперспирализацию путем сшивания образующихся разрывов или разрезания ДНК. Наконец, открыты специальные ферменты, «редактирующие» ДНК, т.е. осуществляющие вырезание и удаление ошибочно включенных нуклеотидов или репарирующие повреждения ДНК, вызванные физическими или химическими факторами (рентгеновское излучение, УФ-лучи, химический мутагенез и др.).
Из клеток животных выделено несколько ДНК-полимераз, и в разных лабораториях они получили различные наименования.
К настоящему времени у эукариот, как и у бактерий (см. ранее), открыто несколько ДНК-полимераз. В репликации ДНК эукариот участвуют два главных типа полимераз – α и δ. Показано, что ДНК-полимераза α состоит из 4 субъединиц и является идентичной по структуре и свойствам во всех клетках млекопитающих, причем одна из субъединиц оказалась наделенной праймазной активностью. Самая крупная субъединица ДНК-полимеразы а (мол. масса 180000) катализирует реакцию полимеризации, преимущественно синтез отстающей цепи ДНК, являясь составной частью праймасомы. ДНК-полимераза δ состоит из 2 субъединиц и преимущественно катализирует синтез ведущей цепи ДНК (см. далее). Открыта также ДНК-полимераза ε, которая в ряде случаев заменяет δ-фермент, в частности при репарации ДНК (исправление нарушений ДНК, вызванных ошибками репликации или повреждающими агентами). Следует отметить, что в эукариотических клетках открыты два белковых фактора репликации, обозначаемых RFA и RFC. Фактор репликации А выполняет функцию белка – связывание одноцепочечной ДНК (наподобие белковых факторов связывания разъединенных цепей ДНК при репликации у Е. coli), фактор С – функцию стабилизатора всего реплика-ционного комплекса.
В генетической инженерии с целью получения белков в достаточных количествах и с заданными свойствами (например, для генотерапии наследственных и соматических болезней) широкое применение получили эндо-нуклеазы рестриктазы, катализирующие расщепление молекулы двух-цепочечной ДНК по специфическим нуклеотидным последовательностям внутри цепи. Рестриктазы узнают определенные 4–7-членные последовательности, вызывая, таким образом, разрывы в определенных сайтах цепи ДНК. При этом образуются не случайные последовательности, а фрагменты ДНК строго определенной структуры с липкими концами (ре-комбинантные ДНК), используемые далее для конструирования гибридных молекул и получения генно-инженерной, биотехнологической продукции (например, инсулина, гормона роста, интерферона, вакцин против вируса гепатита В, СПИДа и др.).
Общий механизм синтеза ДНК. Основываясь на данных о двухспираль-ной антипараллельной структуре, химическом составе ДНК (см. главу 3) и значении «активированной» формы энергии для биосинтеза полимерных молекул, А. Корнберг еще в 1955 г. указал на возможность синтеза ДНК энзиматическим путем в бесклеточной системе в присутствии изолированной из Е. coli ДНК-полимеразы и предшественников дезоксирибонук-леозидтрифосфатов. Реакция, практически осуществленная в 1967 г., сводится к синтезу новой молекулы ДНК:
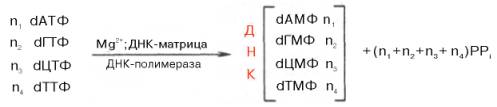
Химический смысл полимеризации состоит в том, что свободная 3'-гидроксильная группа матрицы атакует α-фосфатную группу соответствующего присоединяемого нуклеозидтрифосфата (определяется природой азотистого основания затравки), при этом происходят отщепление остатка пирофосфата и образование фосфодиэфирной связи. Далее свободный 3'-гидроксил вновь присоединенного нуклеотида атакует α-фосфатную группу следующего нуклеозидтрифосфата, и таким путем продолжается процесс полимеризации, идущий в направлении 5'–>3', антипараллельно матрице, оканчивающейся 5'-фосфатом:
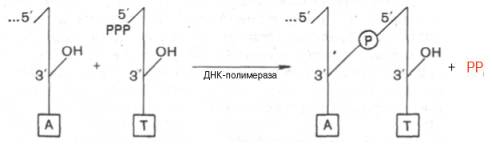
Реакция требует присутствия одноцепочечной ДНК или в крайнем случае небольшого полидезоксирибонуклеотида. В деталях выяснено значение предобразованной ДНК в механизмах действия ДНК-полимераз:
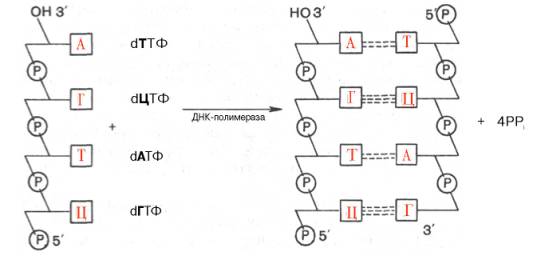
ДНК служит не только затравкой, но и матрицей, на которой фермент комплементарно и антипараллельно синтезирует дочернюю цепь ДНК. Это можно представить в виде схемы:
Были предприняты другие подходы к выяснению механизма поли-меразной реакции. В лаборатории А. Корнберга был открыт фаг (φХ174, содержащий одноцепочечную кольцевую ДНК. Эту молекулу использовали в качестве матрицы в ДНК-полимеразной реакции и получили биологически активную ДНК фага, использовав фермент ДНК-лигазу, обладающую способностью катализировать соединение (сшивку) концов разрывов в молекуле ДНК. Было показано, что в процессе репликации одноцепочечная ДНК фага (φХ174 проходит стадию образования двухцепо-чечной кольцевой ДНК. Применив ряд остроумных подходов, А. Корнберг и сотр. в опытах in vitro создали искусственную молекулу фага φХ174, обладающую способностью поражать (инфицировать) Е. coli, вызывая лизис бактерии. Последовательность событий может быть представлена на схеме, где исходная молекула кольцевой ДНК фага φХ174 обозначена плюсом (+), а вновь синтезируемая молекула – минусом (–) (рис. 13.1). М. Мезельсон и Ф. Сталь показали полуконсервативный механизм репликации ДНК, включающий образование дочерних молекул ДНК, в каждой из которых сохраняется лишь одна родительская цепь (рис. 13.2; 13.3).
Сложность процесса репликации ДНК объясняется тем, что обе цепи реплицируются одновременно, хотя имеют разное направление (5'–>3' и 3'–>5'); кроме того, рост дочерних цепей также должен происходить в противоположных направлениях. Элонгация каждой дочерней цепи может осуществляться только в направлении 5'–>3'. Р. Оказаки высказал предположение, подтвержденное экспериментальными данными, что синтез одной из дочерних цепей осуществляется непрерывно в одном направлении, в то время как синтез другой дочерней цепи происходит прерывисто, путем соединения коротких фрагментов (в честь автора названы фрагментами Оказаки), в свою очередь синтезирующихся в противоположном направлении (рис. 13.4).
Как видно, синтез ведущей цепи ДНК идет всегда в направлении 5'–>3', соответствующем направлению движения репликационной вилки. Сохраняя правило синтеза дочерних молекул ДНК 5'–>3', синтез на второй цепи родительской ДНК идет в направлении, противоположном движению репликационной вилки. В зависимости от типа клетки фрагменты Оказаки имеют разные размеры – от нескольких сот до нескольких тысяч нуклеотидов (150–200 у эукариот и 1000–2000 у бактерий).
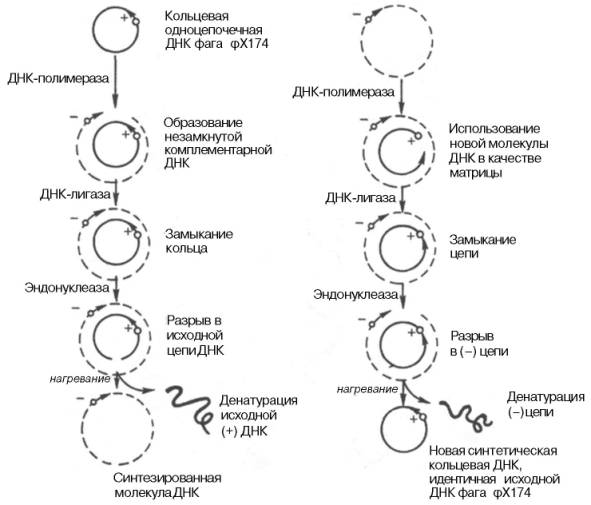
Рис. 13.1. Роль ДНК-полимеразы и ДНК-лигазы в синтезе кольцевой одноцепочеч-ной ДНК фага φХ174.
Получены доказательства, что образование каждого фрагмента Оказаки требует наличия короткого затравочного комплементарного праймера – участка РНК, синтез которого катализируется праймазой. Затем при участии ДНК-полимеразы III синтезируются длинные участки ДНК. РНК-затравки далее вырезаются при участии ДНК-полимеразы I, а свободные места их (бреши) замещаются (достраиваются) комплементарными дез-оксирибонуклеотидами под действием той же ДНК-полимеразы I; наконец, сшивание разъединенных участков отстающей цепи осуществляется при помощи ДНК-лигаз. Подобный механизм челночного синтеза ДНК легко объясняет фактические данные о накоплении коротких фрагментов ДНК у Е. coli во время репликации ДНК.
Особенности репликации ДНК у эукариот. Репликация ДНК у эукариот, по существу аналогичная репликации ДНК у прокариот, имеет ряд особенностей. Например, вместо одной точки репликации в ДНК эукариот имеются специфические точки «начала», так называемые автономно реплицирующие последовательности (около 300 нуклеотидных пар); в дрожжевой клетке таких элементов около 400. Кроме того, скорость движения репликационной вилки у эукариот (примерно 50 нуклеотидов в секунду) почти в 10 раз ниже, чем у E. coli. Для репликации ДНК генома человека из одной-единственной точки с подобной скоростью потребовалось бы более 500 ч; вместо этого репликация генома человека происходит в обоих направлениях и одновременно из множества точек (множество «начал» репликации), вовлекая от 30000 до 330000 пар оснований. Репликация продолжается до тех пор, пока не будут синтезированы две дочерние молекулы ДНК, в каждой из которых содержится одна родительская цепь (см. рис. 13.4). Таким образом, множественность точек «начала» репликации ДНК, вероятнее всего, является общим правилом для всех клеток эукариот.
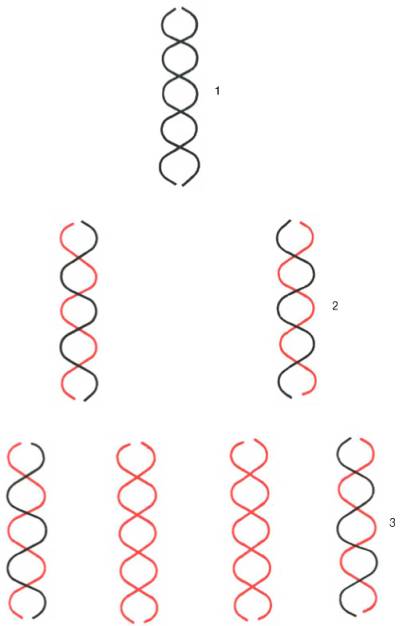
Рис. 13.2. Полуконсервативная репликация ДНК in vitro. Каждая из двух цепей родительской ДНК служит матрицей для синтеза дочерних молекул ДНК. 1 - родительская молекула; 2 -дочерние молекулы (первая генерация); 3 - дочерние молекулы (вторая генерация).
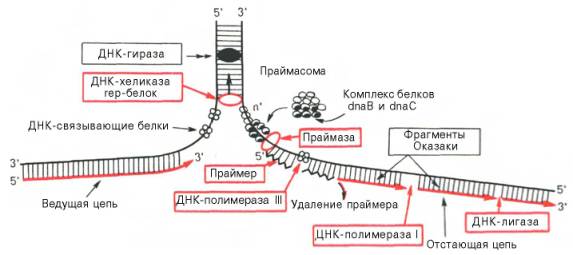
Рис. 13.3. Основные этапы репликации ДНК (схема).
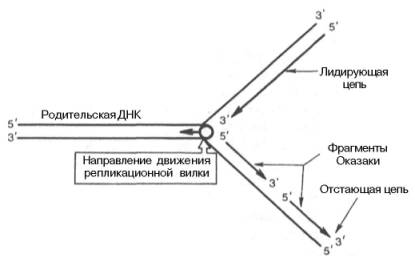
Рис. 13.4. Схематическое изображение непрерывного и прерывистого синтеза цепей ДНК при репликации.
Как было указано, инициация биосинтеза дочерних цепей ДНК требует предварительного синтеза на матрице ДНК необычного затравочного олигорибонуклеотида, названного праймером, со свободной гидроксильной группой у С-3' рибозы. Этот короткий олигорибонуклеотид синтезируется комплементарно на матрице ДНК при участии особого фермента – прай-мазы, наделенной РНК-полимеразной активностью.
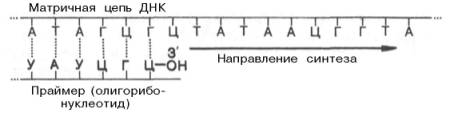
Предполагают, что именно с этой точки концевого 3'-гидроксила рибозы праймера начинается истинный синтез лидирующей дочерней цепи ДНК, комплементарной родительской. Синтез начинается с реакции между 3'-ОН-группой концевого рибонуклеотида праймера и α-фосфатной группой первого дезоксирибонуклеотидтрифосфата в строгом соответствии с комплементарностью родительской цепи ДНК, при этом освобождается пирофосфат. В дальнейшем этот фрагмент РНК, комплементарно присоединенный к новообразованной цепи ДНК, разрушается под действием ДНК-полимеразы I, и возникшая брешь застраивается олигодезоксирибо-нуклеотидом при помощи той же ДНК-полимеразы I. Вполне допустимо предположение, что синтез праймера из олигорибонуклеотида имеет глубокий биологический смысл, поскольку в этом случае могут устраняться ошибки, неизбежно возникающие при инициации репликации ДНК.
Этапы биосинтеза ДНК. Предложен ряд моделей механизма биосинтеза ДНК с участием указанных ранее ферментов и белковых факторов, однако детали некоторых этапов этого синтеза еще не выяснены. Основываясь главным образом на данных, полученных в опытах in vitro, предполагают, что условно механизм синтеза ДНК у Е. coli может быть подразделен на три этапа; инициацию, т.е. начало, элонгацию, т.е. продолжение, и терми-нацию, т.е. завершение (прекращение) синтеза. Каждый из этих этапов требует участия специфических ферментов и белковых факторов.
Этап I – инициация биосинтеза ДНК – является началом синтеза дочерних нуклеотидных цепей; в инициации участвует минимум восемь хорошо изученных и разных ферментов и белков. Первая фаза – это, как указано ранее, ферментативный биосинтез на матрице ДНК необычного затравочного олигорибонуклеотида (праймера) со свободной гидроксиль-ной группой у С-3' рибозы. При инициации к цепям ДНК последовательно присоединяются ДНК-раскручивающие и ДНК-связывающие белки, а затем комплексы ДНК-полимераз и праймаз (см. рис. 13.3). Инициация представляется единственной стадией репликации ДНК, которая весьма тонко и точно регулируется, однако детальные механизмы ее до сих пор не раскрыты и в настоящее время интенсивно исследуются.
Этап II – элонгация синтеза ДНК – включает два кажущихся одинаковыми, но резко различающихся по механизму синтеза лидирующей и отстающей цепей на обеих материнских цепях ДНК. Синтез лидирующей цепи начинается с синтеза праймера (при участии праймазы) у точки начала репликации, затем к праймеру присоединяются дезоксирибонуклеотиды под действием ДНК-полимеразы III; далее синтез протекает непрерывно, следуя шагу репликационной вилки. Синтез отстающей цепи, напротив, протекает в направлении, обратном движению репликационной вилки и начинается фрагментарно. Фрагменты всякий раз синтезируются раздельно, начиная с синтеза праймера, который может переноситься с готового фрагмента при помощи одного из белковых факторов репликации в точку старта биосинтеза последующего фрагмента противоположно направлению синтеза фрагментов. Элонгация завершается отделением олигорибонуклеотидных праймеров, объединением отдельных фрагментов ДНК при помощи ДНК-лигаз и формированием дочерней цепи ДНК. Нельзя исключить, однако, возможности сопряженного и согласованного механизма синтеза лидирующей и отстающей цепей ДНК при участии полимераз и всего комплекса праймасом.
Этап III – терминация синтеза ДНК – наступает, скорее всего, когда исчерпана ДНК-матрица и трансферазные реакции прекращаются. Точность репликации ДНК чрезвычайно высока, возможна одна ошибка на 1010 трансферазных реакций, однако подобная ошибка обычно легко исправляется за счет процессов репарации.
Синтез ДНК на матрице РНК. Выдающимся достижением биохимии нуклеиновых кислот является открытие в составе онковирусов (вирус Раушера и саркомы Рауса) фермента обратной транскриптазы, или ревертазы (РНК-зависимая ДНК-полимераза), катализирующего биосинтез молекулы ДНК на матрице РНК. Накоплены данные о том, что многие РНК-содержащие онкогенные вирусы, получившие наименование онкорнавирусов, содержат ревертазу в составе покровных белков. Фермент открыт также во многих клетках прокариотов и эукариотов, в частности в лейкозных клетках, пролиферирующих тканях, включая эмбриональные ткани. Ревертаза онкорнавирусов содержит ионы Zn2+и активируется катионами Мn2+ и Mg2+ . Предполагают, что синтез ДНК на матрице РНК происходит в 3 этапа. На I этапе фермент ревертаза синтезирует на матрице вирусной РНК комплементарную цепь ДНК, что приводит к формированию гибридной молекулы. Второй этап – разрушение исходной вирусной РНК из комплекса гибридной молекулы под действием РНКазы. Наконец, на III этапе на матрице цепи ДНК комплементарно синтезируются новые цепи ДНК. Ревертазной активностью обладают и ДНК-полимеразы: например, фермент из Е. coli способен катализировать синтез ДНК на матрице рРНК.
Открытие обратной транскриптазы имеет большое значение не только для выяснения закономерностей процесса малигнизации, но и для всей науки о живом, поскольку указывает на возможность передачи наследственной информации от РНК на ДНК, не подчиняясь основному постулату (поток информации идет только в одном направлении):
ДНК –> РНК –> Белок.
В настоящее время можно дополнить эту основную схему передачи генетической информации в живой клетке и представить ее в более полной форме:
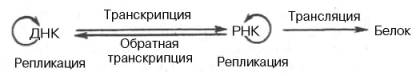
На схеме стрелки вокруг ДНК и РНК указывают на возможность молекул копировать самих себя в живых системах при участии соответствующих ферментов. Как знать, не станем ли мы свидетелями открытия принципиальной возможности поворота стрелки и на следующей стадии – от белка на РНК, что могло происходить на Земле при зарождении первичных живых существ?