Липопротеины
Липопротеины плазмы подразделяются на две группы: белки, связанные с липидами ковалентно, и белки, связанные с липидами нековалентными связями. Липид, ковалентно связанный с липопротеином, служит якорем, с помощью которого белки прикрепляются к мембране. Липопротеины второй группы не имеют строго определенного состава. Они скорее представляют собой агрегаты липидов с белками. Эти липопротеиновые комплексы имеют переменные размеры и состав. В плазме крови они обеспечивают транспорт водонерастворимых липидов.
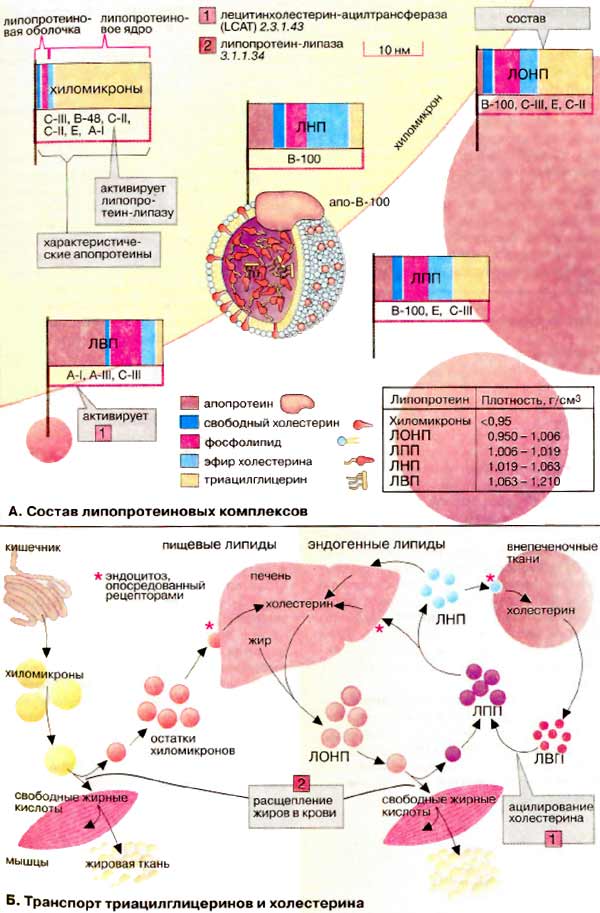
А. Состав липопротеиновых комплексов
Липопротеиновые комплексы представляют собой шаровидные агрегаты, состоящие из ядра, образованного неполярными липидами (триацилглицеринами и ацилхолестеринами), и оболочки толщиной примерно 2 нм, построенной из апопротеинов и амфифильных липидов (фосфолипидов и холестерина). Наружная сторона оболочки полярна, вследствие этого липиды растворимы в плазме. Чем больше липидное ядро, т. е. чем большую часть составляют неполярные липиды, тем меньше плотность липопротеинового комплекса.
Липопротеиновые комплексы делятся на пять групп. Ниже они приведены в порядке уменьшения размера и увеличения плотности: это хиломикроны и остатки хиломикронов, липопротеины очень низкой плотности [ЛОНП (VLDL от англ. very low density lipoproteins)], липопротеины промежуточной плотности [ЛПП (IDL от англ. intermediate density lipoproteins)], липопротеины низкой плотности [ЛНП (LOL от англ. low density lipoproteins)], липопротеины высокой плотности [ЛВП (HDL от англ. high density lipoproteins)]. Липопротеиновые комплексы несут на внешней поверхности характерный апопротеин, который «плавает» на оболочке (здесь в качестве примера ЛНП). Апопротеины играют решающую роль в функционировании липопротеинов: они служат молекулами узнавания для мембранных рецепторов (см. ниже) и необходимыми партнерами для ферментов и белков, которые участвуют в метаболизме и обмене липидов.
Б. Транспорт триацилглицеринов и холестерина
Хиломикроны обеспечивают транспорт пищевых липидов от кишечника к тканям. Хиломикроны образуются в слизистой кишечника и транспортируются в кровь лимфатической системой (см. с. 266). В мышцах и жировой ткани они разрушаются липазой липопротеинов, активирующейся апопротеином С-II. Под действием этого фермента хиломикроны быстро теряют бóльшую часть своих триацилглицеринов. Остатки хиломикронов утилизируются печенью.
ЛОНП, ЛПП и ЛНП тесно связаны между собой. Они транспортируют триацилглицерины, холестерин и фосфолипиды от печени к тканям. ЛОНП образуются в печени и могут превращаться, как и хиломикроны, в ЛПП и ЛНП путем отщепления жирных кислот. Образующиеся ЛНП снабжают холестерином различные ткани организма.
ЛВП возвращают избыточный холестерин, образующийся в тканях, обратно в печень. Во время транспорта холестерин ацилируется жирными кислотами из лецитина. В этом процессе участвует лецитинхолестеринацилтрансфераза (КФ 2.3.1.43). Между ЛВП и ЛОНП также происходит обмен липидами и белками.
Эндоцитоз, опосредованный рецептором. При потреблении холестерина клетки с помощью мембранных рецепторов, узнающих апо-В-100 и апо-Е, связывают ЛНП. «ЛНП-рецепторы» захватывают эти комплексы путем эндоцитоза. Поглощение происходит в так называемых «окаймленных ямках» — областях мембран, у которых внутренняя поверхность выстлана белком клатрином. Клатрин облегчает включение в ямку и содействует отделению везикулы ("окаймленная везикула"). Внутри клетки клатрин отделяется от везикулы и используется повторно. Везикула связывается с лизосомой, которая переваривает ее содержимое (см. рис. 229).